
看得见还是看不见, 这是一个问题!
如果问你, 你现在看到了什么, 你可以不假思索地说, 就是眼前的屏幕嘛. 如果稍微受过一点生理学或者神经科学训练的人也许会说, 我们看到的是这个充满电磁波的世界里, 光学波段辐射被眼球的光学元件折射之后, 落在我们视网膜上的影像.
然而我们不总是能看见落到我们视网膜上的东西. 稍加反思, 我们都会想到自己夏日午后看着窗外出神的时候, 其实并没有看见窗外有什么景色;对着讲台发呆的时候也没有看到黑板上的字是什么;对着击球手飞速击出的棒球同样略过了视网膜, 但是否产生了知觉只有他知道. 简而言之, 就像英语中see(看到)与look(看的动作)不同, 我们看到的东西绝不是落在我们视网膜上的光子所携带的所有信息1——你看不见不可见光也看不到偏振. 那么是什么决定了我们能看到什么以及看到的世界是什么样子呢?
我们可以拍脑门的说, 当我们用心去看就能看到(比如黑板或者窗外), 如果不用心, 那刺激就被我忽视(Neglect)了嘛——的确没错, 这就将我们引导到了视觉注意(Visual Attention)的领地. 不过这篇post中我们先不讨论注意的问题, 而要去关注一些与视觉意识相关的更基本的情况.
什么时候我们看不到 (When we do not see)
首先, 我们将自然环境以及落在视网膜上的光子一概称为物理刺激(Physical stimuli), 我们感觉到的那个世界称为主观感受(Perception)2. 那么刺激与感受不完全匹配的情况其实比比皆是, 下面举几个例子:
- 边缘错觉(Contour Illusion): 有时通过巧妙的几何设计我们会”看到”刺激中并不存在的部分. 例如经典的Kanizsa三角形, 我们会“看到”不存在的三角形边界, 即所谓错觉边界(illusory boundary).

- 运动错觉(Motion Illusion): 有时我们感受到的世界并不是他本来的样子. 如下图(Flash Lag Effect)中如果我们将注意力放在移动的红点上, 则会感受到两点共同出现时红点更靠右. 而事实上两者仅出现在在视频一帧上相同x坐标的位置上, 是纵向对齐的. 因而不对齐的效果是视觉加工的结果. (更有趣的一点是, 如果我们的注意力放在绿点上并一直期待绿点的出现, 那上述不对齐(misalignment)现象就会削弱许多. 因此一些研究者将这一现象归因于我们对视觉场景的期待. )

- 瞬态知觉与视觉掩蔽(Masking): 掩蔽(Masking)翻译得接地气一点就是遮盖, 说白了就是通过一个刺激来影响另一个刺激引起的知觉. 通常是呈现一个极短时间(20-100ms)的暂态(transient)图片, 然后紧接着呈现一个噪声图片或是干扰刺激. 如果不加掩蔽刺激, 我们便能看清图片, 然而由于干扰刺激, 我们没办法识别前一个刺激中的精细信息. 所以这些信息也输入了我们眼中, 然而还没被加工进入意识就被后一个刺激打乱了.

必须要承认视觉系统在加工图像时改变了视觉输入的许多方面, 使得我们感受到的世界与这个真实的世界有所不同. 如果我们意识到的不是我们视网膜上的图像, 那我们能意识到我们脑中神经活动渲染的世界嘛? 这个问题换一种说法讲就是, 我们意识到的视觉世界与哪里的神经活动相关呢, 如果不是视网膜, 那么是大脑中的什么皮层呢? (视网膜听起来还很低级, 而大脑以及皮层则是各种高级功能甚至意识的所在地. 不过皮层看起来很平等, 但一些皮层比另一些更平等. )
怎么找意识的神经相关物?(How to Find Neural Correlate of Visual Consciousness)
研究视觉意识的相关物(统称NCC问题), 首先要区分必要与充分条件.
问什么是视觉意识的必要条件, 我们都知道视网膜, 视神经, 外侧膝状体(LGN), 初级视皮层(V1)都是早期视觉加工的必要通道, 如果损坏了这些神经组织, 显然我们看不到眼前的东西, 自然没有视觉意识. 3 不过, 这些生物结构似乎是必要不充分的. 而要寻找充分条件则是很不同的问题. 特别的, 一些初级视皮层损坏的患者即使报告说看不见眼前的东西, 依然能依据视野中的东西作出正确的判断4, 这一现象被称为盲视(blindsight). 所以V1是否必要, 也存疑问.
寻找视觉意识相关物的一大困难在于, 视皮层的神经元既被外界刺激激活又被内在的心理过程影响. 大多数日常情形下我们意识到的世界与外界刺激本身强烈相关, 于是我们就很难将记录到的神经反应相关于视觉意识本身5, 因为它可能只是低级刺激加工的结果, 并没有被意识到——就像视网膜上神经元的反应. 所以寻找视觉意识的神经相关物的实验关键就是要将视觉刺激与意识分离(dissociate)开来, 让两者不再相关. 比如双眼竞争(Binocular rivalry), 比如模棱两可(Ambiguity)造成的视觉双稳态(Perceptual bi-stability)都是经典的实验范式. 这些范式中外界刺激没有改变, 而内在的知觉变了, 我们能清楚地将神经活动相关到视觉意识上. 而另一些时候, 使用相同的临界刺激(Threshold stimuli), 我们感觉到或是没感觉到同样可以比较出视觉意识带来的不同.
换一个思路, 许多经典的启动效应(priming)与后效(aftereffect)范式也能帮助寻找意识的相关物. 因为, 这些效应就其定义来说, 就是某种没有被觉知到的刺激(阈下刺激), 影响了后续对意识到的刺激的响应. 很多时候这被看做是一种最基本的”潜意识活动”, 或者是基本的学习/记忆行为. 不过现代神经科学的视角下, 这代表了一种在意识之外的神经活动的记忆性 / 历史依赖性——比如一个神经元刚刚被激活之后, 下一次激活的反应会变弱, 这就是一种记忆性. 而如果一些神经元的活动与没被感知到的刺激信息(比如强度太弱的亮点, 过于密的条纹, 无法看清的色彩块)相关, 即包含了比意识表象更多的信息. 那我们也不会认为这些神经元的活动可以被意识到. 因此如果一些神经活动与上述启动效应相关, 那这些活动就不直接与视觉意识相关.
以上的范式大多是在正常人正常意识状态下进行的, 而一些奇特的意识状态如幻觉(Hallucination), 想象(Imaginery), 与做梦也可以用来探究视觉意识, 这些情况下视觉表象和外界世界完全无关, 其内容就可以更加丰富多彩了. 而且正所谓”脑残病人欢乐多”, 通过观察部分脑损伤病人的视觉意识变化, 我们也能将功能定位到一些脑区, 从而找到哪些脑区是必要的, 以及哪些脑区是不充分的.
下面仅举一个经典范式, 来展示下科学家们通常是怎么寻找意识相关物的.
双眼互搏 (Left VS Right)
双眼竞争是很常见的视觉现象, 读者现在就可以尝试: 只需要将手掌放在自己鼻子前面, 让一只眼睛看到手心, 一只眼睛看到手背, 如果你把注意力聚焦在近处(手掌附近而非远处景象/屏幕), 那么你很快就会感觉到手心和手背的感知在交替着主导着你的视野. 经典研究表明每只眼睛主导的平均时长大约10s左右, 呈现一个右偏的分布.
在2005年的实验中4, 实验者向被试左眼输入红色条纹, 右眼输入蓝色垂直方向条纹, 让被试随时报告主观感知到的图像, 实验者就可以通过这些报告来标记时间轴, 并训练分类器来解码被试看到的图像. 更进一步直接记录被试正常情况下观看红色条纹与蓝色条纹的响应, 也可以训练分类器来识别被试到底看到了哪种图像.
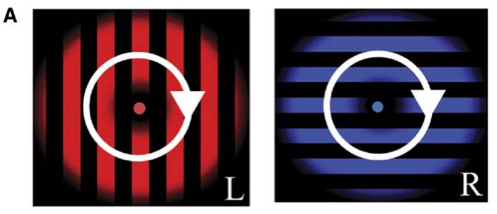
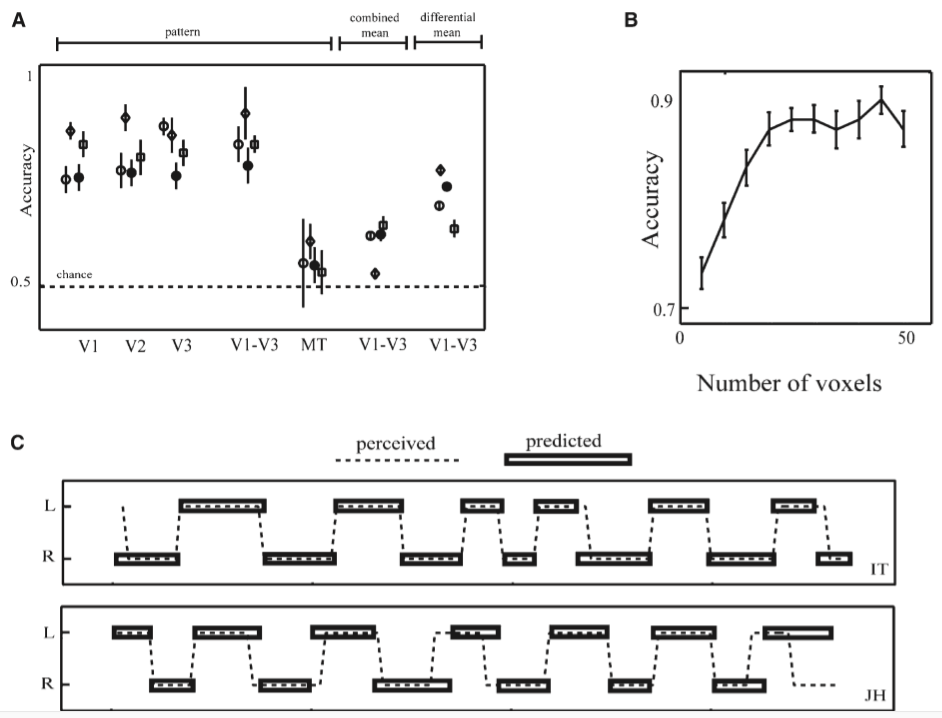
下图显示了根据不同脑区的活动模式训练的分类器的成绩, 以及随着体素(Voxel)数量6增加而成绩的提高. 下图中用V1-V3的活动模式训练分类器都可以取得不错的成绩, 也就是说即使在fMRI的测量精度下依然可以从V1-V3早期视觉皮层读出不同眼睛的输入信息, 也就是当时的视觉表象.
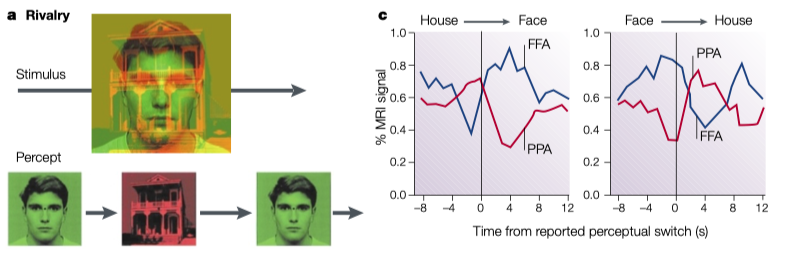
上述实验的刺激可能过于初级, 我们同样可以使用复杂图像刺激输入双眼, (如房子和人脸)7 . 如同我们预期的, 在编码复杂刺激的脑区(如FFA对人脸, PPA对地点, 环境有选择性)同样可以观察到随着视觉表象的交替, FFA与PPA的激活也随之交替.
不过这两个实验不太令人满意, 总感觉这一范式有些初级. 在人身上, 双眼信息在V1才第一次汇聚到同一个神经元上. 所以可以想象由于某种局部的神经计算(比如相互抑制(mutual inhibition), 适应(adaptation)以及与LGN之间的反馈动力学(feedback dynamics)), 两眼输入交替主导着初级视皮层的活动. 于是下游的神经活动也被两种刺激交替主导. 但我们并不能肯定V1的活动模式是否在更精细的意义上与我们的视觉表象对应. 因此在人类的双眼竞争的现象中, 初级视皮层编码了哪只眼睛的图像成为我们的视觉表象, 但我们依然不知道初级视皮层否足以产生我们的视觉意识.
我们能意识到V1的神经活动吗(Are we aware of V1 activity?)
这个问题也是1995年Christopher Koch与Crick一篇著名猜想文章的标题(Are we aware of neural activity in primary visual cortex?).
这个问题的表述就可以进行一番概念讨论, 显然我们并不能意识到V1某个神经元在每时每刻发放了多少个动作电位——否则也就不需要电生理仪器做什么了, 只要让研究生回家记录自己的神经元发放了多少个脉冲就行. 显见, 包括神经活动在内的许多生物细节信息对主体是不透明的. 因而当我们说意识到V1的神经活动, 并不是指这个.
在更一般的意义上, 意识内容, 物理刺激以及神经活动之间的关系显然不是同一(identical)的关系(你意识到的红色并不等同于某个波长, 你听到的音调不等同于某个振动频率, 也不等同于某组神经活动模式. ) 但他们会有相关关系(correlation)8. 如果相关这个词比较”难听”, 我们可以更定量化, 说他们有互信息(mutual information). 比如不同朝向的条纹可以激活V1中不同偏好(tuning)的神经元, 因而我们可以讲V1神经元群体编码(encode)了朝向信息, 同样我感知到的不同朝向条纹与不同神经元集群的发放, 也是相关关系. 更一般的, 只要神经活动会随着主观感知 / 刺激变化(modulation), 这种互信息关系就存在, 我们就可以谈”神经活动编码了xxx”. 当然, 除了刺激内容, 还有时间上的相关性, 当一组刺激出现 总是伴随着一组神经活动模式, 以及一种特别的感知内容(允许有时间上的延迟), 那么他们就有一种相关性.
这种相关性是一般科学视角下讨论意识与神经活动关系的概念根基9. 所以这里我们实际追问的是V1的神经活动与我们的视觉意识在什么方面是相关的. 粗粗看来V1神经活动的许多方面与视觉意识紧密相关, 比如它编码了视野中条纹的局部朝向; 比如由于视野到视皮层的连续映射(Retinotopy), 神经元激活的位置也代表了刺激在视野中的位置.
然而, 在2002年的一篇综述10里也概括了V1神经活动与视觉表象在许多方面的不同(dissociation), 我在这里整理如下.
- 解剖上, V1对前额叶, 运动前皮层(prefrontal premotor)没有直接投射, 因而传统上认为无法直接提供信息给决策中枢.
- 心理物理学上, V1常常中包含了超出视觉意识的信息.
- 如果(通过特制眼镜)视觉刺激仅从两眼中的一个射入, 你是无法分辨刺激是从哪只眼睛进来的. 然而V1包含着对左眼或右眼有偏好的细胞组成的(Ocular Dominance Stripe). 因而包含了刺激来源的信息.
- 极其密集的条纹(high frequency grating), 对人眼看起来就像一片均匀的颜色一般, 然而依然会有与条纹的方向相关的视觉后效(aftereffect)——这说明条纹的方向信息一定被编码在某处, 但我们没有意识到它. 一般认为朝向选择性最早出现在初级视皮层V1, 所以这也说明V1编码了一些无法被觉察到的信息.
- 电生理记录也表明单神经元的活动模式与视觉意识流很不同.
- 眨眼(blinking)通常不会打断视觉意识流, 但V1的神经元在眨眼时则近乎被关闭了——正如视网膜上的图像被眼帘完全关闭了一般. 在眨眼期间V1神经元发放率甚至比外界刺激关闭时的发放率还低11. 这种V1活动的不连续性告诉我们, V1神经元有更短的时间常数, 对外界反应更快, 而我们的视觉意识也许与更高层时间常数更长的皮层有关.
- 微眼动(microsaccade), 是一种我们通常不会觉知到的眼球的微小跳跃——就在此时此刻, 你在阅读时也会有频繁的眼动行为, 然而你对世界的知觉是连续, 稳定的, 尽管落在视网膜上的图像随着眼动会大幅度变化. 而当有刺激划过感受野, V1神经元会随着微眼动有推迟的激活现象, 这与主观上感受不到微眼动有些冲突12.
- 当色彩条纹高速转动时, 我们便无法分清其中每一种颜色, 然而神经元依然可以对其中的单一颜色响应——这也说明视皮层的单神经元中包含着比知觉中更多的信息. (就像落在视网膜上的图像也包含比知觉更多的信息. )
- 视觉表象可以独立于V1活动改变: 在猴子身上做双眼竞争任务, V1的单神经元更多的反映了输入刺激的稳定性质, 而非猴子的视觉表象.
- 与此不同的是人身上就能从V1的fMRI活动模式上提取出视觉表象信息…也许猴子与人的V1功能不尽相同
- 对于一般人, 常出现梦境的快速眼动睡眠(REM)期间, V1活动被抑制, 但不妨碍视觉表象的出现. 失去部分或全部V1的病人, 依然能做视觉梦境, 而双侧颞枕叶切除的病人才会完全失去做视觉梦境的能力.
- 单侧顶叶脑损伤(parietal damage)病人对于损伤对侧的视觉刺激经常难以觉察. 然而那些没被觉察到的对侧视觉刺激在V1激发起的活动模式与觉察到时很像, 因此, 至少在这些病人身上, V1的活动本身并不能支持对刺激的视觉意识.
总而言之, V1单神经元的神经活动与视觉表象的活动模式有许多不一致, 包含的信息也超出了视觉意识的范围, 同时并不足够解释/支撑视觉意识——所以在这篇10综述中作者的结论就是我们意识不到V1的神经活动, 就像我们意识不到视网膜上细胞的活动一样. 而基于一些类似但更复杂的证据作者认为, 高级视觉区域与我们的视觉意识能更好的关联起来. 也许V1中神经元集群的活动模式也能与视觉表象关联, 也许是某种更复杂的脑区间交互作用模式才带来了意识, 但这些都需要更好更大规模的神经记录手段才能了解.
总结(Summary)
以上数据大多是在正常意识状态下进行的, 视觉感知的内容也比较固定(看到了vs没看到, 左眼图像vs右眼图像), 而一些更奇妙的状态下我们的视觉意识与外界世界完全分离, 这时视觉表象与神经的关联就更加有趣了, 我将在下一篇中介绍.
限于篇幅, 本文需要到此打住了! 意识是极其复杂的科学问题, 这里我引入了这个问题的一个小小的入手点——视觉意识, 以及前人是怎样试图定位这一意识的神经相关物的. 总的来说, 这篇综述10认为与视觉意识与高级视觉皮层中的神经活动更相关, 而初级视觉皮层的神经活动则被认为不太会被意识到. 这些结论至今还没有完全建立, 而且也没有一个被所有人接受的意识理论, 因而将神经活动和意识现象对应起来就更加困难了.
功能定位通常是神经研究的第一步: 把功能局域化到特定的脑区, 就能拿起放大镜仔细研究这一脑区的独特属性, 追问更深层的问题: 她有什么样的投射模式(projection pattern)? 什么特殊的神经元类型(Cell type)? 特殊的局部网络(microcircuitry)? 特殊的胞外环境? 特殊的蛋白表达以及细胞内机理? (说不定真的有量子机制在某个特殊的脑区起作用产生了意识也未可知…) 然后或许能找到意识更微观的机制.
但另一种可能性是, 我们再努力也无法将意识定位到一个脑区中. 现在观测到的相关性也许只是在我们有限的神经活动记录精度下的结果, 一些更精微的活动模式被磨掉了, 因而看不到与一些脑区的相关性. 当我们真的掌握了全脑的神经活动(如对斑马鱼), 也许会发现意识状态与全脑的神经活动模式都有关系. 那时, 我们该怎么办? 也许对意识重要的不是空间, 而是时间——如同步的神经活动(Synchrony), 高频神经震荡(Gamma Oscillation), 或是不同脑区之间的信号传递(Recurrent / Feedback)10. 而意识状态也许是这些神经变动的结果, 而无法被保存在凝固的状态里. 在一个真正的网络/相互作用系统中, 我们无法将一个全局的性质划归到任何一个节点上. 如果是这样, 那这种真正的涌现(emergence)现象就更激动人心了. 让我们拭目以待.
-
特别的, 心理学也会区分Sensation and perception, 感觉与知觉, 一个更初级更接近物理刺激, 一个更加接近我们的主观感受. ↩
-
心灵哲学中也会称之为Qualia即感受质, 即我们所感觉到的世界的样子. 然而即使扫一下上述参考文献的长度就可以知道关于这一概念的哲学争论是多么的丰富, 此处我们指出与Qualia的关联, 但无意在哲学上下什么结论. ↩
-
如果读者看过17年的电影敦刻尔克的话, 一定记得其中有个情节是被拯救的盟军飞行员阻止船长把船开回去救人, 争吵中把船长的儿子撞下了舷梯磕伤了后脑勺, 然后他说自己什么也看不到了. 让我猜的话很可能是后脑勺对应的枕叶(Occipital lobe)的初级视皮层被破坏导致什么也看不见, 即使他前端的视网膜与视神经看起来完好无损. ↩
-
Haynes, J.-D., & Rees, G. (2005). Predicting the stream of consciousness from activity in human visual cortex. Current Biology, 15(14), 1301–1307.6 ↩ ↩2
-
统计的语言说, 如果有两个强相关的回归因素(regressor), 那么仅通过回归就很难区分出这两个变量谁的贡献更大. ↩
-
这里涉及一个fMRI的分析方法, 所谓多体素模式识别(Multi Voxel Pattern Analysis). 对于机器学习来说, 有一个活动模式, 能从中解码出一些信息并不神奇. 不过有趣之处在于它对于fMRI竟然也能用. 众所周知, fMRI的每个体素代表了大约1mm^3量级的空间信息, 对于神经元来说已经是数万个细胞组成的集群了. 而由于BOLD信号直接来源于血管而非神经元, 它测量到的空间信息可能更粗. 在这种粗粒度下, 竟然还能解码出诸如刺激的方向, 色彩等信息, 很大程度上是因为, 每个体素是对携带不同信息神经元的一种不均匀的加权平均, 而不同体素的偏好有些许不同, 如果综合大量体素来训练分类器, 而非直接平均, 就能重新得到不同类神经元的信息. ↩
-
Tong, F., Nakayama, K., Vaughan, J. T., & Kanwisher, N. (1998). Binocular Rivalry and Visual Awareness in Human Extrastriate Cortex. Neuron, 21(4), 753–759. https://doi.org/10.1016/S0896-6273(00)80592-961 ↩
-
在一些哲学讨论中, 这种关系也被称为表征(representation)关系, 但这个概念的精细刻画还需要与哲学系的小伙伴探讨下. ↩
-
事实上哲学家对意识的神经相关物有大量概念讨论, 我这里无法穷尽, 希望了解心灵哲学的哲学系朋友们可以补充一下. ↩
-
Rees, G., Kreiman, G., & Koch, C. (2002). Neural correlates of consciousness in humans. Nature Reviews Neuroscience, 3(4), 261–270. https://doi.org/10.1038/nrn7830 ↩ ↩2 ↩3 ↩4
-
Gawne, T. J., & Martin, J. M. (2000). Activity of Primate V1 Cortical Neurons During Blinks. Journal of Neurophysiology, 84(5), 2691–2694. https://doi.org/10.1152/jn.2000.84.5.2691 ↩
-
Martinez-Conde, S., Macknik, S. L., & Hubel, D. H. (2000). Microsaccadic eye movements and firing of single cells in the striate cortex of macaque monkeys. Retrieved from http://neurosci.nature.com ↩