
先天与后天 (Nature or Nurture)
首先让我们从一个更大的背景开始.
长久以来, 从哲学家, 心理学家到神经科学家, 许多探索人类本性的研究者都会讨论所谓先天与后天的问题. 哲学家们追问的是人性, 即人作为人的规定性(definitive property)是先天的还是后天的: 在某个讨论中是说, 人到底先天上是有内容, 被规定的, 还是就是一块白板呢1; 在心理学的语境中讨论的是行为或心理规律的来源 — 这些规律到底是先天具有, 还是后天被培养, 教导, 学习(learning), 适应(adaptation)才逐步形成的呢. 在物质层面的神经机制上, 我们的神经系统也被两种过程决定, 一部分是生理的发育过程(development)2, 一部分是后天的学习, 记忆, 适应(learning, memory, adaptation)变化. 前者很大程度被基因决定 (genetically-encoded), 是分子机器的运转诱导下精确的环路组装3; 后者则由输入的环境刺激, 以及神经系统自身的可塑性(neural plasticity)决定, 是高度个性化与”文化”相关的.4 用常见的神经-计算机类比粗略来讲, 被基因决定的前者像是事前烧好的电路板, 即硬件(hardware); 可以学习适应的后者就像是软件(software), 可以在较快时间尺度内修改.
而就像计算机一般, 神经系统在大致相同的硬件基础上, 根据输入经验、训练, 甚至内在状态的不同, 可以运行非常不同的程序, 执行极其不同的任务. 就像平时你为了快乐而读一本书, 和哲学课上为了写论文构造某个论证或寻找某个证据而读一本书, 结果会很不同, 后者会让你更敏锐的挑选那些相关信息并深度加工. 而在我们平时训练猴子或者老鼠做不同的行为实验时, 根据不同的给予奖赏的方式, 猴子与老鼠可以很快的改变输入/输出模式, 来获取更多的奖励.5 这种能力有时被叫做情境依赖的计算功能(context dependent computation). 而对于我们人类, 我们甚至可以直接用语言, 改变他人的内在状态, 让人们去做什么或者不做什么. (比如, 助教对你说 “再不交报告你这门课就要挂科啦! ““啊啊啊我赶紧回去写TAT”) 分析起来, 就是通过给予对方信息, 改变对方对不同行为结果优劣的期望(Reward expectation of different behavior outcome)的估计, 从而改变其自发行为输出.
铺垫了这么多学习和经验对神经系统的影响, 好像我们大部分的行为都是这样的, 人生下来啥也不会, 都需要学习. 但对于大部分动物, 他们生下来就几乎准备好了做各种事情, 那么这些所谓先天行为, (innate behavior) 所对应的神经基础就应该是被遗传物质编码好, 并在神经发育时稳定形成的. 那么什么样的行为会被所谓硬件编码(hardware coded)进基因中呢? 对于人, 我们依旧很不清楚, 但对于动物, 我们清楚的知道一些重要的行为通过其环路结构被编码进基因里.
果蝇的嗅觉系统(Drosophilia’s Olfactory System)
在进入正式的结果前, 再铺垫一点关于果蝇的嗅觉系统的背景知识.
对应于我们的鼻子, 果蝇的嗅觉感受器(Olfactory Receptor)主要分布在两处器官上: 触角(antennae)与maxillary palps. 在这些感受器官上分布着不同的感受神经元(Olfactory Receptor Neuron, ORN), 感受神经元膜上的受体蛋白(ligand-gated metabotropic channels)结合一些气味分子, 引起一系列细胞内信号分子(如cAMP)的产生, 最终使膜去极化并产生动作电位传递到下游神经元(Action Potential, Spike). 每个感受神经元上的受体蛋白不同, 能接收的气味分子也不同, 因而感受神经元激活的模式(activation pattern)就可以告诉他的大脑它到底闻到了什么气味.

果蝇的触角上有大约1300个感受神经元, 他们会用将轴突投射到Antennal lobe上的43个嗅球上(Glomerulus5), 相同类型(即接收相似气味信息)的感受神经元汇聚到几个同类嗅球上, 进行信号汇聚.
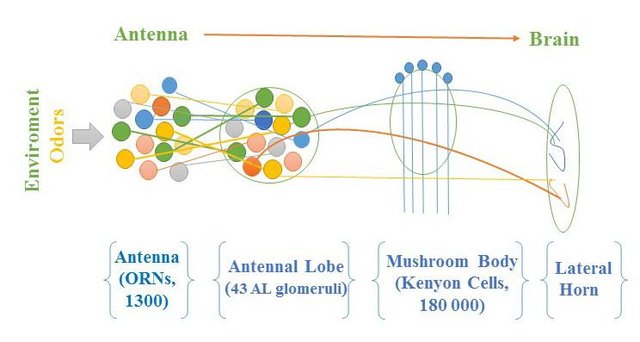
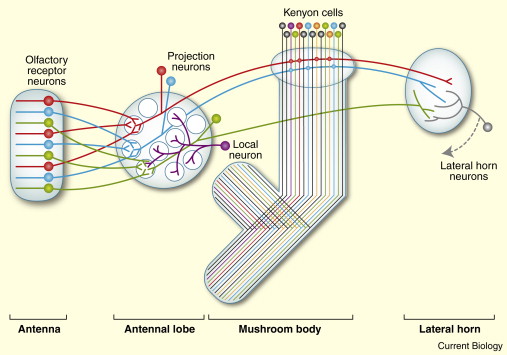
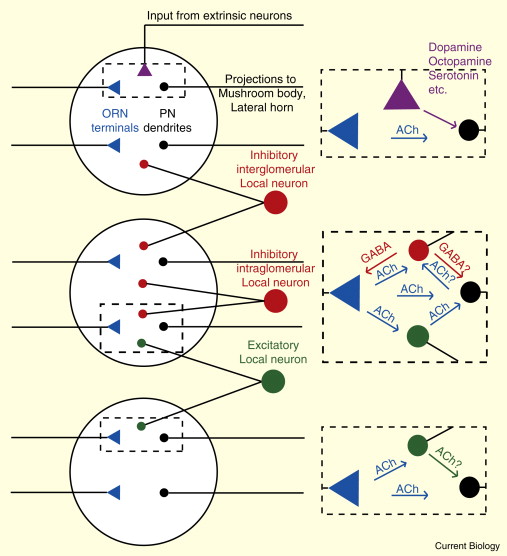
与嗅球相关的有两类不同的神经元. 投射神经元(Projection Neuron, PN)将自己的树凸伸到嗅球里, 接收感受神经元(ORN)的输出, 并将信号接力至蘑菇体(Mushroom Body, MB 因为长得真的很像蘑菇= =)与侧角(Lateral Horn,LH)上. 嗅球中的另一些神经元(Local Neuron)则不会长程投射, 而是负责局部的计算: 比如在嗅球内或两个嗅球间相互抑制或者激活. 改变嗅觉信息的表征. (见上下图6)
再往下走, 蘑菇体(MB)与侧角(LH)是嗅球的主要下游区域, 他们负责着很不同的功能, 蘑菇体与嗅觉学习(associative olfactory learning)很有关, 近几年的工作也发现了诸多气味的效力(valence, 带来奖赏/惩罚, 是吸引的还是排斥的)是如何在学习过程中被写入和读出蘑菇体的Kenyon Cell (KC)网络的. 而侧角与行为输出直接相关, 即使蘑菇体被损坏, 侧角也可以介导基本的行为输出.
有了嗅觉系统的基本图像, 这次介绍的神经回路就好理解多了.
果蝇的气味回避行为(Neural Circuit for Avoidance in Drosophilia)
有一种叫Geosmin的常见气味分子(闻起来就是土味…), 是一些常见微生物的代谢产物. 但极少计量就能够削弱果醋对果蝇的吸引力. 而果醋正是果蝇最喜欢的食物信号, 所以Geosmin自然是果蝇很”讨厌”的气味了. 那么这种强烈的排斥行为的神经基础是什么呢?
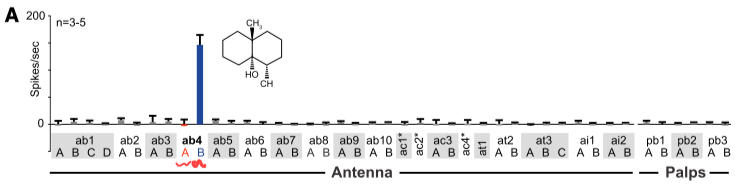
马普所的一群研究者在2012年做了很漂亮的成果7. 他们测量了所有触角上以及口器上的嗅觉神经元(ORN), 发现只有其中一类(ab4B)对Geosmin有强烈的反应; 而更令人惊奇的是他们对自然界中的自然气味(水果, 食物, 粪便等)作了化学成分分解后, 依次刺激该神经元, 在这样的大范围筛选下, 也发现也几乎只有Geosmin一种分子有极强的响应(下图). 这种强选择性(Selectivity)在嗅觉系统里极其罕见了. 果蝇身上另一种与之可比的强选择性受体就是对异性的气味信号分子的接收神经元.
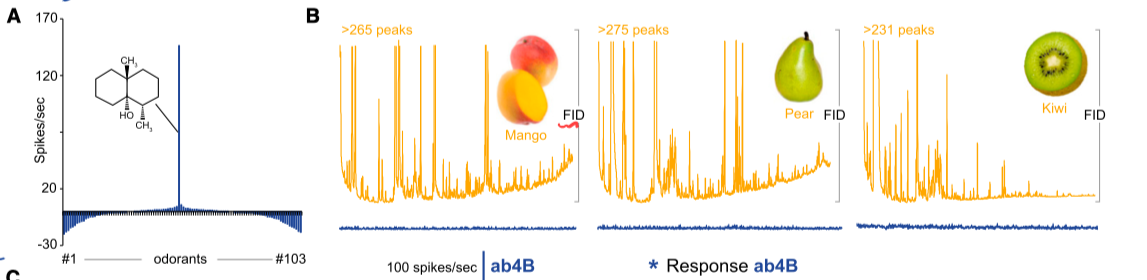
更令人惊讶的是这类神经元的下游, ab4B神经元几乎只投射到Antennal Lobe的一个嗅球(DA2)上的一类投射神经元DA2, 这一类DA2神经元同样对Geosmin有极强的选择性! 而他们发现, 只要激活DA2投射神经元, 就能激发果蝇的回避行为; 而反过来如果完全阻断了ab4B感受神经元到DA2神经元的信号传递, 则回避行为完全不会出现. 因此可以说Geosmin引起的回避行为完完全全是通过DA2神经元兴奋引起的了.
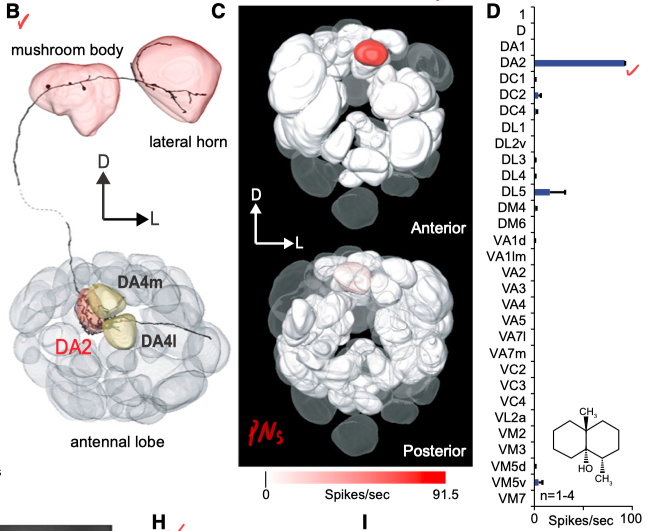
这一神经通路的发现非常惊人, 为什么呢? 因为工程上, 用单一一条神经线路感受仅仅一种气味分子, 即所谓标记线编码(labeled line code)是非常不划算的设计—-想想看触角上总共只有几十种不同的感受神经元, PN总共也只有40余种, 但自然界有成千上万种气味, 不可能被这样一一编码, 因而通常气味是通过感觉神经元特定的共同激活模式来组合编码(Combinatoric coding)的. 而分出一条PN来单独负责(dedicate)一种分子, 就会减少总体能感知到的气味的类型, 缩小嗅觉信道容量. 不过另一方面, 标记线编码也像十万火急的鸡毛信一样, 可以毫无疑问(unambiguously)传递这一种分子的信息, 不用解码, 直接引起下游的输出. 这一逻辑说明, 这条线上(Geosmin-ab4B-DA2)传输的信息应该带有十分重要, 甚至生死攸关的价值8! 但Geosmin为何重要呢, Geosmin本身并不是毒素, 也不影响果蝇的生活, 那么果蝇为什么要特意分出一条神经电缆来感知这一种物质呢?
为了探究这个生态价值的问题, 研究者将目光转向了自然界中产生Geosmin的微生物(e.g. S. coelicolor, P. expansum), 他们发现, 这些微生物对于果蝇的幼虫是致命的, 在含有这些微生物的培养基上, 果蝇幼虫活不了五天! 这样看来能直接影响后代存活与否, 可以说是十分强大的选择力量了: 具有对这种气味的感觉的与回避模式的果蝇可以极大地提高下一代存活率, 因而这种基因和相对应的神经环路被选择出来也就很不奇怪了.
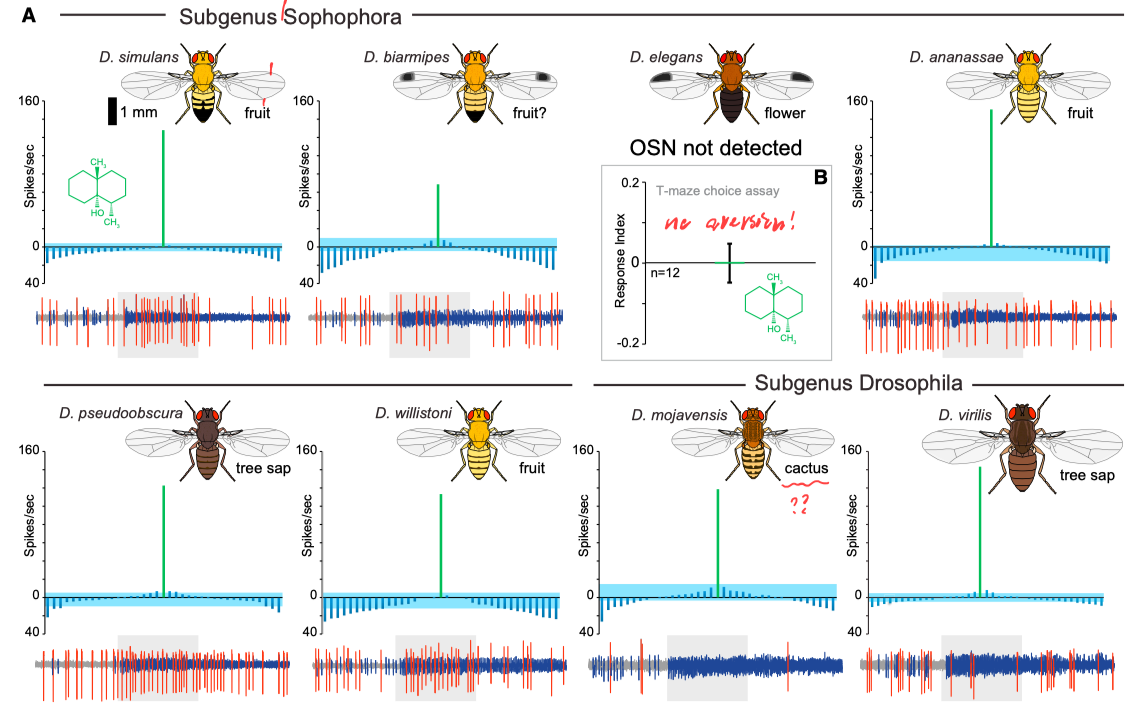
既然这一感觉信号有着如此重要的繁殖价值, 那么很可能这一结构在进化长河中很早就被选择出来了! 事实上的确如此, 研究者检验了其他果蝇品种(下图), 发现了他们的触角上都有相似的一类神经元, 只对Geosmin有极强的选择性, 而且都具有对Geosmin的回避行为. 基于此, 研究者估计这一气味感觉通路是果蝇种(genus Drosophila)的一种根本特点, 在历史中已然存在了四千万年.
有趣的是, 果蝇对这些微生物的感知, 仅仅是通过Geosmin这一种气味信号. 当研究者通过基因工程破坏了微生物产生Geosmin的酶, 果蝇就完全无法分辨这种”悄无声息”的有害微生物, 于是会无差别的在含有他们的培养基上产卵, 然后后代也会因此会死掉. R.I.P. 可以说这是一条非常纯粹但是脆弱的感觉通道了= =
进一步探索(Going Deeper into Circuit)
(前面的故事基本就讲完了, 这部分是最近的后续研究, 有些technical, 如果对学术部分兴趣不大的读者可以跳到最后一部分. )
说来在今年新出的一篇BioRxiv文章中9, 马普所的研究者们更是一路追踪下去, 揪出了Geosmin响应通路更下游的神经环路.
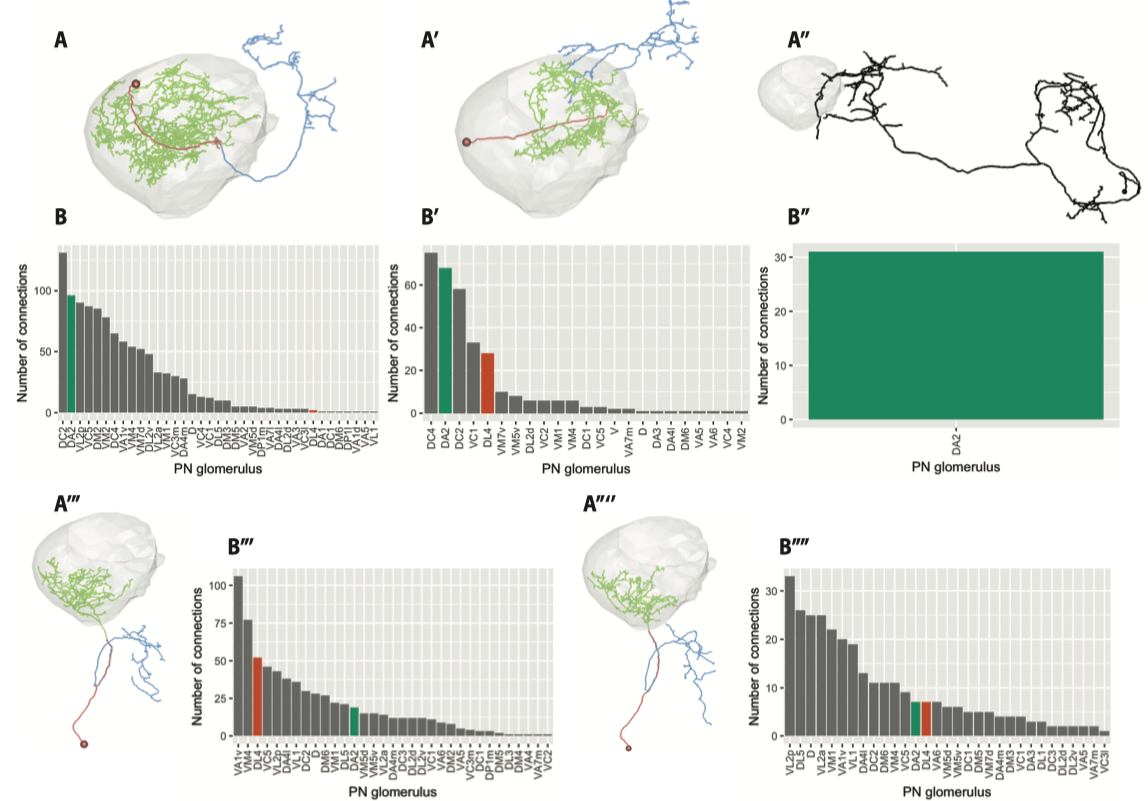
为了追踪DA2投射神经元的下游通路, 首先要判断大方向, 投射神经元(PN)产生行为到底是经过蘑菇体(MB)还是侧角(LH)呢? 他们阻断了蘑菇体的神经元的输出(chemical silencing), 发现并不影响回避行为, 与先前知识吻合, 于是大致排除了蘑菇体的作用. 接下来他们通过电子显微镜与Confocal成像进行显微解剖, 在3D重构的神经元中寻找受到DA2投射的侧角神经元(LN). 有趣的是他们发现接收DA2神经元信号的目标数量远超想象, 他们采样到的151个神经元. 换言之, 这股信息流在侧角LN处发散成了百条溪流走向不同的通路. 而对神经形态做分类后, 他们发现大多是传出神经元(Lateral Horn Output Neuron), 其次是局部神经元(Lateral Horn Local Neuron).
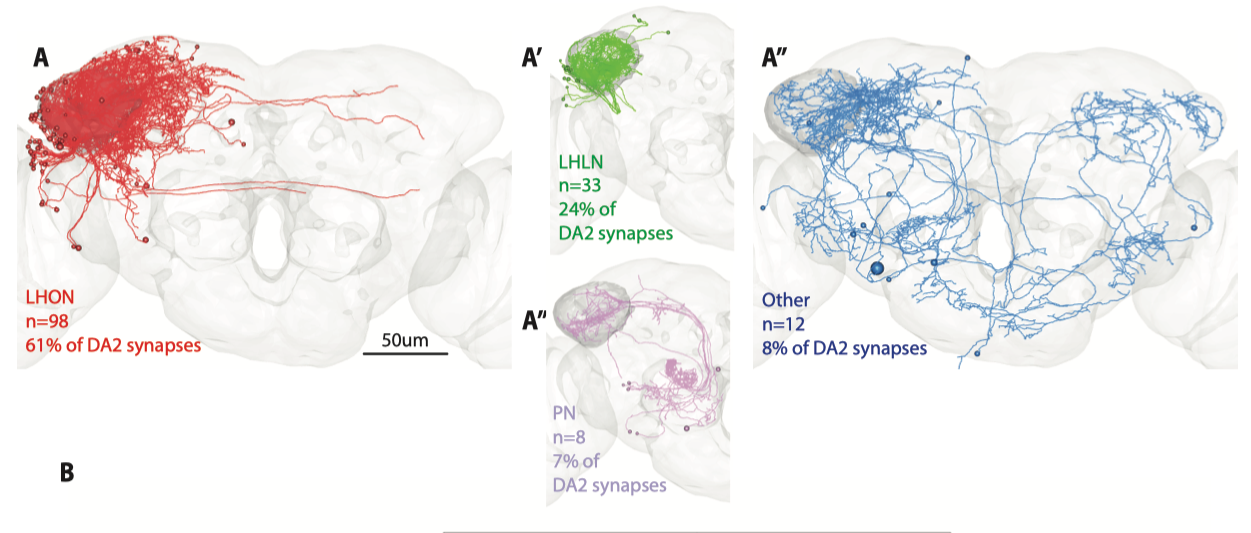
而这样多的下游神经元难以逐个详尽研究, 他们挑选了数个有代表性的LN神经元进行进一步分析, 发现多种果蝇厌恶的气味(包括Geosmin)会汇聚到几个LN神经元上, (见下表PN神经元与LH神经元的投射矩阵) 或许说明这些神经元可以整合不同气味的回避属性(编码果蝇有多厌恶这种味道), 并引起下游一般的回避行为.
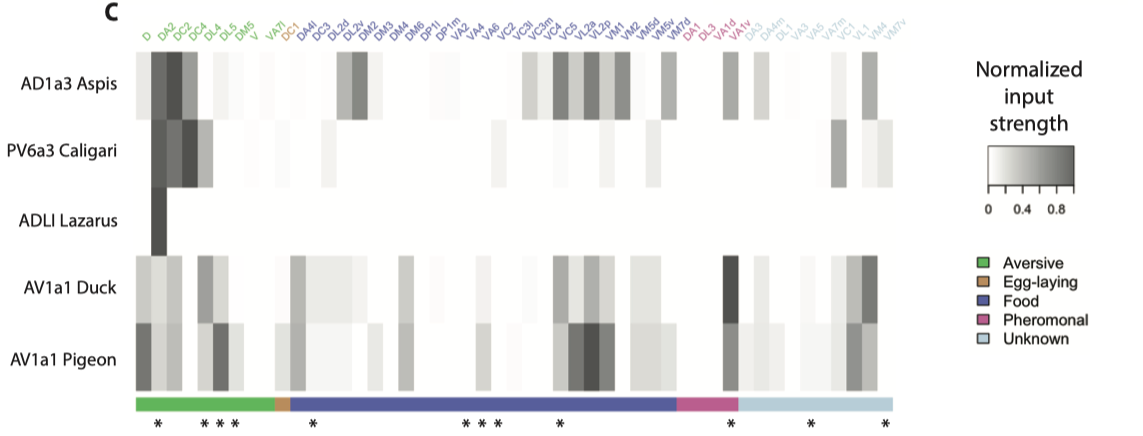
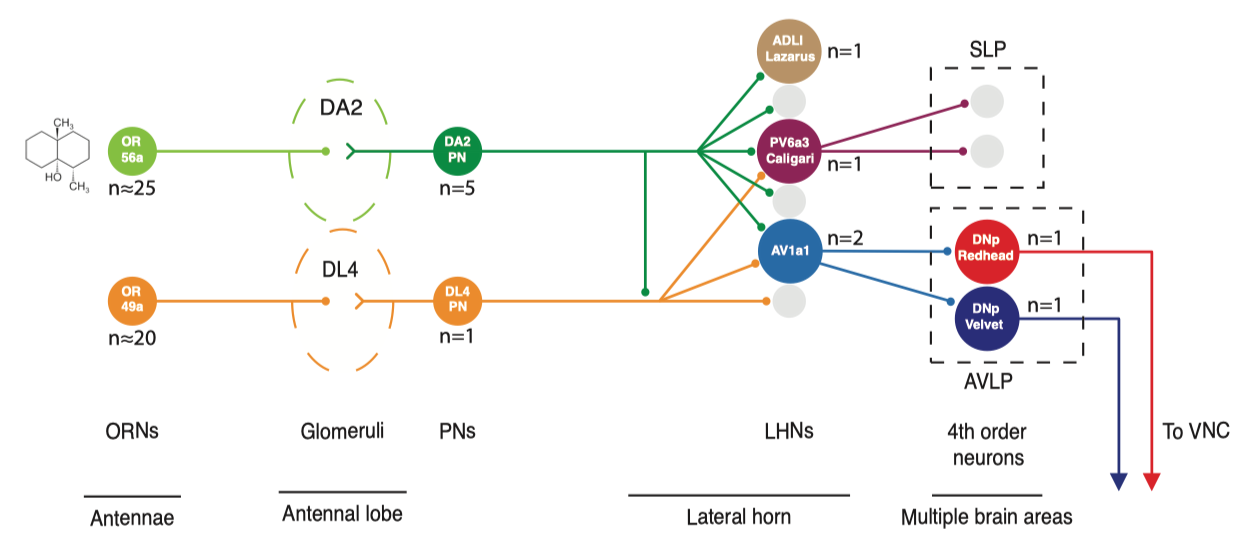
在侧角神经元中, 他们通过复杂的筛选找到了每类神经元对应的光遗传标记, 并发现一类神经元(Av1a1, 下图中的A’’’, A’’'’代号pigeon, duck)对于Geosmin回避行为是充分且必要的—-激活即可引起回避行为, 而阻断其传出即可关闭回避行为. 可以说他们又找到了这个通路在侧角(Lateral Horn)中的一个关键节点(Av1a1).
若从侧角输出神经元(LNON)再进一步追踪下游信号, 由于目标太多难以完整进行, 他们选取了两三个神经元进行追踪. 特别的他们发现, 刚才提到的Av1a1神经元下游投射到DNp Redhead 与 DNp Velvet神经元, 而这两种神经元直接投射至腹侧神经索(Ventral Nerve Cord, VNC), 也就是直接控制肌肉的线路. 因而在这个通路中, 从感觉神经元到肌肉控制最短只需要经过4个突触传递, 就可以将气味信号输入转换成了行为输出. 可以说是非常直接的脑回路了! (上述回路结果被概括在下面的网络图示中, 大体是前馈网络, 只有少许的发散与汇聚)
学术后记(Academic Reflection)
读过这几篇文章7,9,10 写完了这次的post有这样几点感想.
如同上一个post中介绍的小动物的听觉系统选频功能的研究. 本文研究的也是更简单的生命体中的感觉系统的构造. 这次的感觉环路叫环路(circuit)似乎都有点名不副实, 因为他完全就是个前馈通路(feedforward network)… 而一般的前馈通路还有发散与汇聚(Convergence and divergence), 并非一一对应, 但是对ORN->PN的通路, 神经传递甚至是一条线连过去的! 因而前文中7使用的诸多实验技巧才会如此漂亮的展现出其结果: 阻断通路中一环, 下游就完全没有输出; 激活通路中的一环, 下游的输出又出现了. 这样”纯粹”的通路可以说是十分罕见的, 微生物只通过这一种代谢产物Geosmin被觉察, 而果蝇也只通过一种感受神经元(ab4B ORN)以及一类投射神经元(DA2 PN)来传输. 对于神经科学家来说大大简化了问题, 但对果蝇来说这种简单的通路也是脆弱的, 由于果蝇是通过Geosmin分子这单一信道一种感受神经元探查有害微生物, 那么一旦微生物学会不产生Geosmin, 那他们对果蝇就是隐形的! 而这时果蝇也会因为在错误的地方产卵而减少繁殖能力.
另外在 技术上, 数字显微解剖(in silico micro-anatomy)对于环路研究(特别是果蝇这类小生物)很有帮助. 由于神经元本身有着极其复杂(而美丽)的三维结构, 为了观察其结构, 追踪其轴突树凸的走向, 寻找突触, 单纯的染色切片是很不足够的. 电镜或者confocal显微镜与计算3D重构则是一种十分有效的工具. 但单纯的解剖结构又是不足够的, 还是需要功能上的调控(抑制与激活(silencing and activation))来验证结构是work的, 以及重要的.
结构与功能的关系, 前面提到由于这里网络结构, 以及响应模式都出乎意料的简单, 因而从结构出发我们直接就能理解其功能—-神经的连接就意味着信号的传递, 而且信号表征的就是Geosmin这一种气味分子, 信号的结果就是回避行为, 输入与输出都没有疑义. 简直可以说是”过脑子”的最简单的感觉刺激-行为响应映射了. 而如若在更中枢的系统(大脑皮层)做回路研究, 就很难有这样清晰的解读.
后一篇工作震惊到我的一点是做连接组追踪, 以及 理解连接组数据的困难, 我本以为对于果蝇这样小的脑子可以通过计算解剖学轻易的拿到连接图谱, 然后理解从输入到输出的整个网络. 然而看到这片文章才意识到技术上的困难, 他们仅仅是从投射神经元PN努力向下追踪了一层, 就绝望的发现由于下游神经元太多, 很难完全的追踪下去11, 只能挑选几个代表再追下一层. 而且追踪了连接关系, 想去提取其中principle还是很难, 部分原因是连接组研究的个体数量很少, 不知道能否推广, 也无法下很强的结论. 即便在如此简单的情境中(网络被基因决定十分模式化, 前馈结构, 输入输出都可以理解, 很少冗余, 突触传递次数很少), 去理解一个神经网络的逻辑(Principle of network connectivity)还如此困难, 可以想见如果要直接理解皮层中的连接组(Connectomics of Neocortex)逻辑会多么的抓狂.
哲学联想(Philosophical Speculation)
所谓诚其意者,毋自欺也。如恶恶臭,如好好色,此之谓自慊。
–大学
让我们重新回到更广的思维图景中. 朱子在四书章句集注中, 对《大学》这句话的解释12是, 要使修行者爱好善行, 厌恶恶行, 就如同爱好美好的颜色, 或者厌恶恶臭的气味一般, “以自快足于己”, 只因为这样做对自身的满足.
很有趣的是《大学》中举的这两个例子几乎对应于果蝇嗅觉系统相关的两种硬件编码(hardware coded)的行为: 闻到异性的气味而被吸引, 或者是闻到伤害指示性的气味而回避. 这些行为几乎就是将感受信号直接转换成行为(甚至只通过4个突触!!)13. 因此十分直接, 不会被干扰. 这种倾向几乎保持到了人类身上, 因而《大学》也就以这种行为作为自发、无需训练、无需反思、不得不如此(necessary)的喜好或者厌恶的代表, 可以说是非常有道理了.
这种低等动物身上的研究之所以会让人们惊讶, 也是因为人身上很少有完全的硬件编码行为. 人似乎总能通过训练, 通过内在更高层的控制(overwrite)来改变行为模式, 去控制动物的”本能”, 去改变简单的刺激输出映射, 去克服自然对我们的编程: 我们不会见到”好色”就不可抑制的想冲上去发生性行为, 我们不会因为恶臭而拒绝给亲人端屎擦尿.
回到前面的《大学》里的那句话, 对于人来说“恶恶臭, 好好色”的自然倾向是存在的, 但人们不像果蝇一般会直接由刺激导向行为. 我们有着更大的脑子, 有着更复杂的高级调控, 因而除了这些自然倾向, 人们有更多可以提供内在奖赏与外在奖赏的方式, 道德体系, 社会关系等等. 某种意义上人能够能够抑制, 甚至能够改变自己恶恶臭, 好好色的性质, 也是人的神经系统更发达的结果呢~
-
更有争议的例子比如, 黑人同学的成绩不好, 是因为黑人先天就笨基因就差呢, 还是他们更多的来自更低教育水平的家庭环境? ↩
-
很有意思的是, 发育生物学(developmental biology)与发展心理学(developmental psychology)都叫的development, 但所处理的时间尺度与阶段很不同(从中文翻译的不同也可见一斑). 前者处理的更多是被基因与分子机器决定的统一的过程, 后者更多的涉及包括后天学习与适应的过程, 并存在个体差异. ↩
-
因此一些与神经环路组装(circuit assembly)相关的蛋白质对应基因的变异, 直接会引起神经环路的”硬件问题”, 然后导向神经系统功能的障碍. 如自闭症Autism就是较常见的神经发育疾病, 而一种发病机制就是Shank基因的病变, 这一基因对应着突触后的中心支架蛋白Shank蛋白. 因而一个蛋白的损坏可以引起整个回路组装的失调. ↩
-
一个最近听说的例子是, 猴子(非人灵长类)动物脑袋里视觉皮层中对面孔响应的神经元区块(Face Patch)是需要在猴子成长过程中多多接触面孔的视觉刺激才会发育而成的. ↩
-
中文翻译得不清楚, Glomerulus的拉丁词含义是小线团(ball of thread), 可以说很贴切了. 肾小球也叫这个. ↩ ↩2
-
图片来源 https://www.researchgate.net/publication/279743049_OLFACTORY_SYSTEM_IN_DROSOPHILA/figures ↩
-
Stensmyr, M. C., Dweck, H. K., Farhan, A., Ibba, I., Strutz, A., Mukunda, L., … & Wicher, D. (2012). A conserved dedicated olfactory circuit for detecting harmful microbes in Drosophila. Cell, 151(6), 1345-1357.这篇paper里的工作极其漂亮而且工作量极大, 纯粹是为了基础科学的兴趣, 老师评价是在美国的科研体制下不可能做出来的工作了. ↩ ↩2 ↩3
-
之前读过也做过感觉系统最大化信道容量(maximize information content/Channel capacity)的研究, 这回的内容会让我进一步思考最大化信息的意义. 对于生物体, 信息与信息的价值是不同的, 大多数生物并不在乎完整的表征(represent)我们周围的世界—-而且那也是不可能的. 他们要选择性的接收或提取出对自己重要的信息! (就像人眼能且只能看到所谓”可见光”也许也是因为这一波段是阳光辐射最强的波段, 也能提供关于我们感兴趣的日间世界最多的信息.) 而一些并不重要的信息丢掉也无所谓. 所以之后如果再做信息最大化可以考虑加入不同信息价值的权重, 再去最大化. 这件事理论上怎么做也是很有趣. @ 理论研究者们 ↩
-
Huoviala, P., Dolan, M. J., Love, F., Frechter, S., Roberts, R. J., Mitrevica, Z., … & Cornwall, H. (2018). Neural circuit basis of aversive odour processing in Drosophila from sensory input to descending output. bioRxiv, 394403. ↩ ↩2
-
Mansourian, S., Corcoran, J., Enjin, A., Löfstedt, C., Dacke, M., & Stensmyr, M. C. (2016). Fecal-derived phenol induces egg-laying aversion in Drosophila. Current Biology, 26(20), 2762-2769. ↩
-
原文是 “The considerable divergence of the geosmin processing pathway at the 2nd to 3rd order level means that within the scope of this work we are unable to follow all of the connected LHNs deeper into the brain. Instead we chose to focus our efforts on a small number of LHONs we judged to be of special interest.” 可以说十分沮丧了. ↩
-
“言欲自修者知为善以去其恶,则当实用其力,而禁止其自欺。使其恶恶则如恶恶臭,好善则如好好色,皆务决去,而求必得之,以自快足于己,不可徒苟且以殉外而为人也。” https://ctext.org/text.pl?node=89505&if=en&show=meta&remap=gb ↩
-
一个有趣的问题是(food for thought), 如果某些行为是完全硬件编码的, 感觉信号如此直接就可以导向某种行为结果, 那动物真的感受到了这种外界信号了嘛? 还是像条件反射一般, 有了某种信号就直接有了行为. 动物根本不会觉察到自己接收了这种感觉, 而只能在自己已经做出行为之后再”意识到”自己感觉到了什么. @ Philosophers ↩